RAD51 protège contre la réparation mutagène des cassures double-brin de l'ADN
Le maintien de la stabilité du génome est essentiel et, en particulier, les cassures double-brin de l’ADN (CDB) qui sont des lésions très toxiques pour les cellules, doivent être réparées. Mais une réparation inadéquate peut générer des altérations du patrimoine génétique. Des scientifiques de l'Institut Cochin (CNRS/Université Paris Cité/Inserm) décrivent un nouveau rôle de RAD51, protéine-clé de la Recombinaison Homologue (RH), un des mécanismes de réparation des CDB : La seule fixation de RAD51 sur l’ADN - indépendamment de son rôle catalytique actif dans la RH - empêche l’ADN de devenir un substrat pour les autres mécanismes mutagènes de réparation. Ces résultats sont publiés dans la revue Nucleic Acids Research.
Le maintien de la stabilité du génome est essentiel à l’homéostasie cellulaire. L’instabilité génétique conduit à une prédisposition au cancer, au vieillissement accéléré et à des pathologies inflammatoires et neuro-dégénératives. Les cassures double-brin de l’ADN (CDB) sont une lésion des plus toxiques pour une cellule. Elles peuvent être générées par des sources exogènes (exposition aux radiations, molécules chimiques ...) ou endogènes (radicaux libres, perturbation de la réplication du génome). Les CDB doivent nécessairement être réparées pour éviter la mort de la cellule. Plusieurs mécanismes de réparation existent dont certains sont conservateurs de l’information génétique mais dont d’autres génèrent des modifications du patrimoine génétique.
Le choix du mécanisme approprié en fonction de la nature de la lésion, de sa localisation, ou de l’état de la cellule est décisif pour la stabilité du génome. Cette sélection s’organise en deux étapes : Un premier choix entre la protection des extrémités de l’ADN pour favoriser leur ligature directe (NHEJ - Non Homologous End Joining) ou la résection, c’est-à-dire la dégradation partielle des extrémités pour générer des zones d'ADN simple brin (ADNsb). Ces ADNsb sont des intermédiaires nécessaires pour plusieurs systèmes de réparation : i) des mécanismes mutagènes comme l’appariement de zones simple brin complémentaires (Single Strand Annealing, SSA) ou la ligature des extrémités après résection (Alternative End Joining, A-EJ), ii) la Recombinaison Homologue (RH) qui est le seul mécanisme conservatif du patrimoine génétique intervenant après la résection. La protéine clé de la RH, RAD51 (conservée dans toutes les espèces vivantes) se fixe sur l’ADNsb et scanne l’ADN intact dans le génome pour recherche une séquence qui sera utilisée comme matrice pour réparer la CDB, maintenant ainsi la stabilité du génome.
Les scientifiques ont voulu comprendre les mécanismes par lesquels RAD51 empêche les réparations mutagènes post-résection. Pour cela ils ont travaillé avec des cellules humaines en culture dans le génome desquelles ils ont introduit des systèmes rapporteurs permettant de mesurer la réparation par chacun des mécanismes NHEJ, SSA, A-EJ et RH. Les CDB sont introduites par l’expression d’une méganucléase (I-Sce1) entraînant des coupures ciblées dans le génome.
Les chercheurs ont observé que la déplétion de RAD51 (ou BRCA2 qui permet la fixation de RAD51 à l’ADN ; BRCA2 est fréquemment muté dans les cancers du sein ou de l’ovaire héréditaires) stimule à la fois le SSA et l’A-EJ, mais pas le NHEJ, validant le modèle en deux étapes. Ils ont ensuite utilisé trois formes mutantes "dominantes négatives" de RAD51 (les DN-RAD51) qui répriment la RH et stimulent SSA et A-EJ. Dans les cellules vivantes, ces trois DN-RAD51 ne se fixent pas efficacement sur la chromatine endommagée et empêchent la fixation de la protéine RAD51 endogène. Ces trois DN-RAD51 sont altérés sur la fonction d’hydrolyse de l’ATP de RAD51, ce qui a permis dans un premier temps de conclure que l’hydrolyse de l’ATP par RAD51 était nécessaire à sa fixation sur l’ADN endommagé, in vivo.
Ils ont en parallèle utilisé un quatrième DN-RAD51 qui inhibe également la RH mais ne stimule pas SSA et A-EJ. Contrairement aux trois premiers DN-RAD51, celui-ci se fixe efficacement sur la chromatine endommagée. Par conséquent, la promotion de la RH est dissociable de l’inhibition des mécanismes de réparation mutagènes SSA et A-EJ et cette inhibition est uniquement due à la fixation de RAD51 à l’ADN. Enfin, les auteurs montrent que la fixation de RAD51 à l’ADN ne régule pas l’efficacité de la résection des DSB, mais empêche l’appariement des zones d’ADN simple brin complémentaires qui en résultent, ce qui est une étape nécessaire du SSA et de l’A-EJ.
Ces travaux ont donc permis de montrer que RAD51 contrôle la sélection de la voie de réparation des CDB : il préserve l'intégrité du génome en protégeant les cassures des mécanismes de réparation non conservateurs, ceci en se fixant sur l'ADNsb et indépendamment de la promotion de RH.
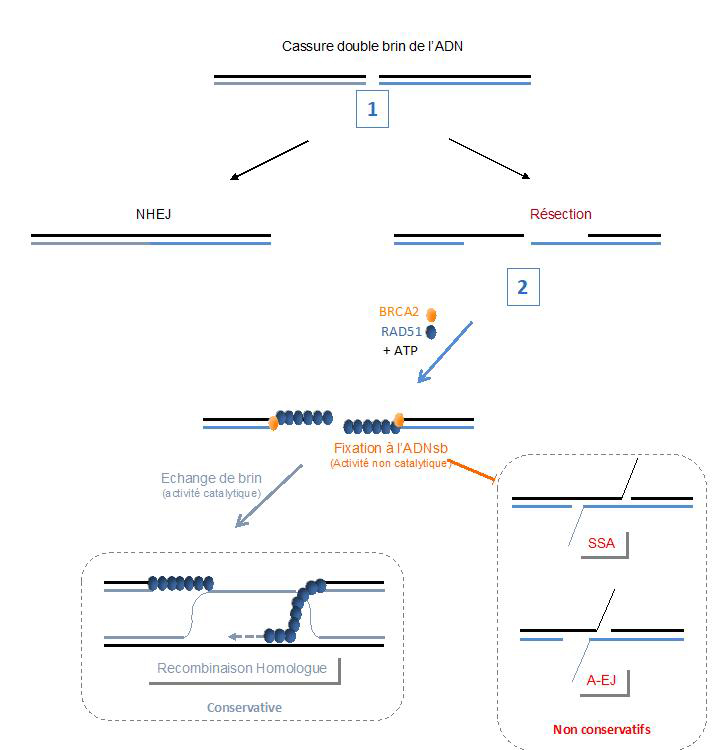
Figure : Les rôles de RAD51 dans la protection contre la réparation mutagène des cassures double brin de l'ADN. 1. Le NHEJ est en concurrence avec la résection sur les DSB. 2. Après résection, BRCA2 charge RAD51 sur l’ADN simple brin. Cette étape est dépendante de l’hydrolyse de l’ATP par RAD51. L'occupation de l'ADNsb par RAD51 promeut la recombinaison homologue (de manière catalytique) et inhibe l’appariement de séquences complémentaires, qui pourrait être dépendante de RAD52, requise pour SSA et A-EJ non conservateurs. Le blocage de l’appariement de zones complémentaires par RAD51 ne nécessite pas d'activité d'échange de brins.